
文献解读
代谢物如何调控表观遗传
绘谱君最近从代谢物经典调控模式(Nat Rev Mol Cell Biol | 小分子代谢物调控模式知多少)和代谢物非经典调控模式-蛋白质翻译后修饰(小分子代谢物非经典调控模式(蛋白篇))分别进行了整理和总结。今天,绘谱君从代谢物-表观遗传角度为大家整理代谢物如何发挥作用。
生物体的细胞含有几乎相同基因组DNA,而核小体是DNA和组蛋白形成染色质的基本结构单位。每个核小体由147bp DNA缠绕组蛋白八聚体(H2A、H2B、H3和H4)形成。细胞通过代谢动态修饰特定核苷酸或氨基酸残基上的DNA/RNA和组蛋白,这些修饰通过多种机制单独或协同地影响各种基因组相关过程,如DNA复制和修复或转录,以及通过改变染色质局部和全局结构。DNA在胞嘧啶和腺嘌呤残基处可以被甲基化,而RNA可以发生甲基化和乙酰化等修饰。组蛋白N端尾部进行化学修饰,包括甲基化、乙酰化、酰化、羟基化、糖基化等(小分子代谢物非经典调控模式(蛋白篇))。染色质修饰可以分为酶促修饰和非酶促修饰。非酶促修饰是通过组蛋白和DNA共价内聚到代谢产生的亲电部分而发生,是某些细胞异常特征,但其功能仍然知之甚少。为了简单起见,我们将这些修饰统称为表观遗传修饰,它们共同形成表观基因组,并与基因调控以及许多生理和病理过程有关,每一种修饰都来自于代谢的中间产物。
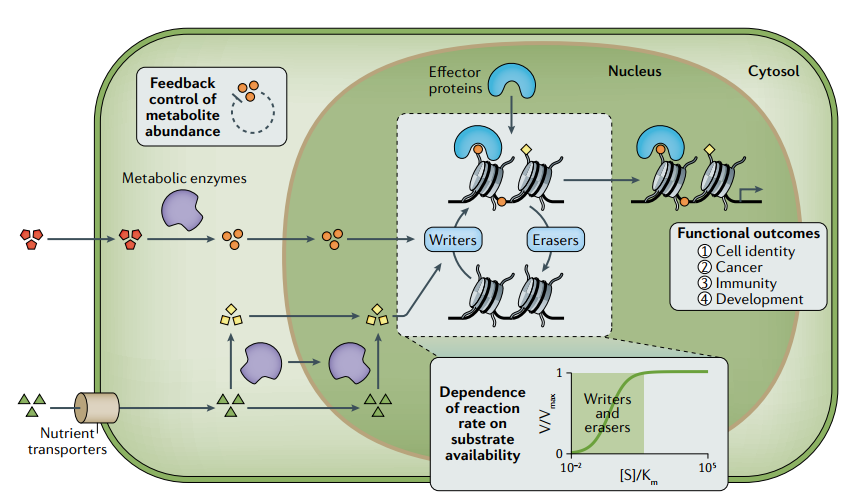
图1. 表观遗传学代谢调控机制简图
代谢与表观遗传之间的联系
由于代谢途径的复杂性,完全理解代谢-表观遗传上下调控作用关系还是存在挑战性,但一些普遍原则是这些修饰和调控的基础。
(1) 表观遗传修饰对代谢活动波动的响应能力是不同染色质修饰酶内在热力学和动力学参数不同的结果。大多数修饰的添加和去除是由酶(即“书写者”和“擦除者”)催化,这些酶利用代谢物作为底物或辅因子(即染色质修饰代谢物)。使用生理浓度接近或低于酶内在Km和Kd值的代谢物的染色质修饰酶比使用过量代谢物的染色质修饰酶更容易受到代谢途径改变的影响。因此,这种特性使代谢波动能够影响某些染色质修饰酶活性,并调节特定表观遗传修饰水平,代谢物可利用性和Km值的差异可能决定表观遗传修饰对代谢改变的相对敏感性。
(2) 另一方面,染色质修饰也可以通过非酶方式发生,在这种情况下,虽然动力学和热力学性质没有很好地表征,但它们在一定程度上受到质量作用定律影响。
(3) 存在几个额外机制使代谢有效和精确的调节酶催化染色质修饰。参与合成染色质修饰代谢物如乙酰辅酶A和S-腺苷蛋氨酸(SAM)的代谢酶,可以定位于细胞核并与核小体和染色质修饰酶相互作用,以便在特定基因组位点上有效地产生代谢物。而染色质修饰代谢物(如SAM)水平又受多种因素调控。这些机制为控制代谢物水平和染色质感知细胞内代谢状态提供了途径。
(4) 表观遗传修饰通过各种机制影响转录程序。所有这些结果都可能受到表观基因组代谢调控的潜在影响。此外,染色质相分离是否受代谢调节仍是一个悬而未决的问题,但研究发现表明,细胞内染色质相分离的能力可能受到代谢物衍生的表观遗传修饰调节,因此可能对细胞代谢敏感。此外,染色质相分离是否也导致代谢物的定位以及在特定相中染色质修饰反应的激活或抑制尚不清楚,但它可能作为精确控制局部代谢物水平和染色质修饰的额外的一种机制。
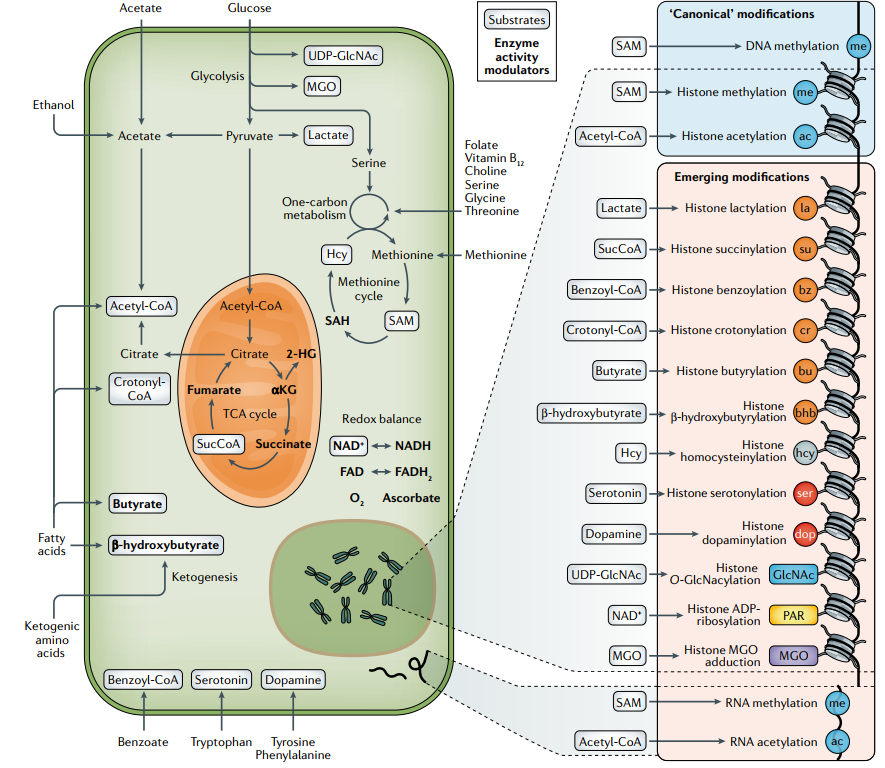
图2. 染色质修饰代谢物的代谢途径及修饰类型
DNA修饰:哺乳动物中存在四种不同DNA修饰,包括胞嘧啶残基5-碳甲基化,产生CpG二核苷酸中的5-甲基胞嘧啶(5mC);5mC氧化为5-羟甲基胞嘧啶(5hmC),进一步氧化为5-甲酰基胞嘧啶(5fC),最终氧化为5-羧基胞嘧啶(5caC)。目前研究最多的是甲基化,这种修饰源于必需氨基酸蛋氨酸(Met)代谢,而在哺乳动物中,蛋氨酸几乎完全从饮食中获得。Met摄取后转化为甲基供体代谢物SAM,SAM随后被用作DNA甲基转移酶的底物,产生S-腺苷型同型半胱氨酸(SAH),其竞争性地抑制DNA甲基转移酶。
DNA去甲基化酶(TETs)TET家族和组蛋白去甲基化酶(包括JmjC结构域去甲基化酶(JHDMs)和胺氧化酶(LSDs)催化DNA甲基化去除。TETs和JHDMs属于α-酮戊二酸(αKG)依赖的双加氧酶,以αKG和氧为底物。这些酶可以被代谢衍生的αKG结构类似物抑制,包括琥珀酸盐、富马酸盐和肿瘤代谢物2-羟基戊二酸盐(2-HG)。TETs和JHDMs的催化作用由抗坏血酸激活,也需要Fe(Ⅱ)作为辅助因子,而LSD组蛋白去甲基化酶依赖于黄素腺嘌呤二核苷酸(FAD)。产生或消耗这些分子的任何代谢途径发生改变如谷氨酰胺缺乏、铁缺乏或线粒体酶(包括IDH1/2、琥珀酸脱氢酶(SDH)和富马酸水合酶(FH))突变,能够通过影响上述代谢物来调节DNA去甲基化酶的活性,从而重塑甲基化。
RNA修饰:目前越来越多研究证实代谢如何影响RNA生物学,如代谢途径和代谢物直接控制翻译机制的选择性和效率,以及RNA转录后修饰来微调蛋白质合成。葡萄糖或氧气减少会导致编码细胞生长和增殖的mRNA翻译抑制,而编码生存并帮助细胞适应新环境的mRNA翻译增加。细胞内NAD+/NADH稳态也可对翻译进行调节。目前已经发现170多种RNA修饰,普遍存在于各种编码和非编码RNA中。然而,直到近十年,RNA修饰的功能(前mRNA剪接、核输出、翻译、稳定和降解等)才逐渐得到认可,尤其是N6-甲基腺苷(m6A)及其生物功能。tRNA修饰以数量最多而闻名,平均每个分子有13个修饰。对于核糖体RNA(rRNA),RNA修饰不可或缺,因为没有假尿苷和2′-O-甲基化,rRNA的生物合成会被中断。长非编码RNA(lncRNA)修饰主要是甲基核苷酸衍生物,包括m6A和m5C。人类小核RNA(snRNA)含有2′-O-甲基、假尿苷和碱基甲基化,参与RNA剪接反应。目前相对研究最多的RNA修饰为甲基化和乙酰化,如N6甲基腺苷(m6A)、N6,2′-O-二甲基腺苷(m6Am)、N1甲基腺苷(m1A)、5-甲基胞嘧啶(m5C)、N4乙酰胞嘧啶(ac4C)、N7甲基鸟苷(m7G)。
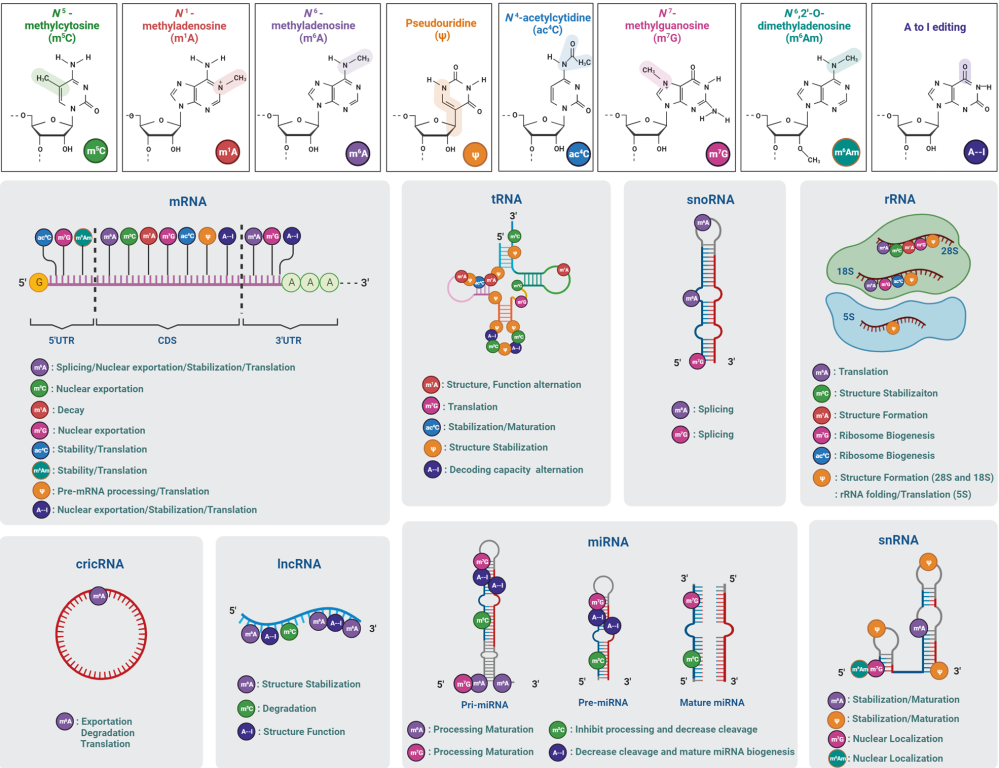
图3. 八种RNA修饰的化学结构、分布和分子功能
组蛋白修饰:组蛋白修饰对染色质结构、DNA复制和修复、转录等动态调节至关重要。除了乙酰化和甲基化修饰外,研究还发现了几种组蛋白标记,包括丙酰化、丁基化、巴豆酰化、2-羟基异丁基化、丙二酰化和琥珀酰化。关于组蛋白修饰(如巴丁酰化和2-羟基异丁基化)的初步研究表明,它们对染色质功能的影响不同于赖氨酸乙酰化。(详细介绍可参考:小分子代谢物非经典调控模式(蛋白篇))
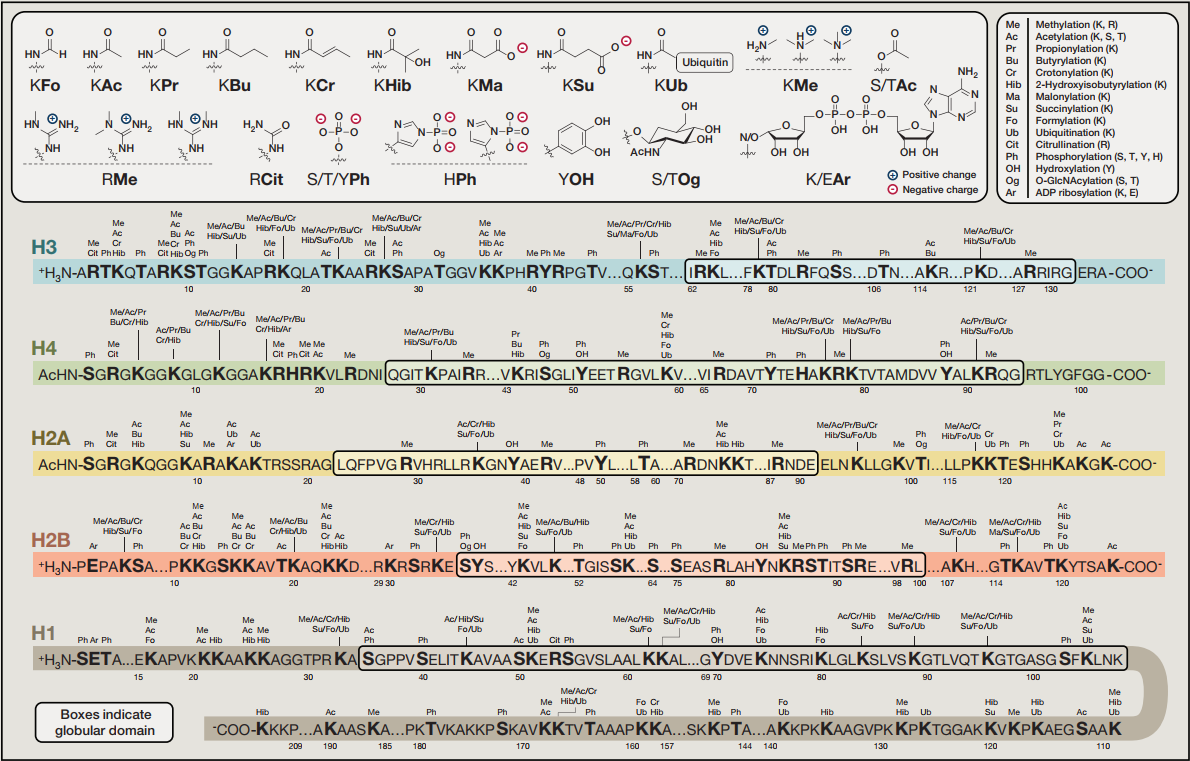
图4. 组蛋白修饰简图
小结
代谢和基因表达是所有生物体必不可少的两个基本生物过程,它们相互调节,维持体内平衡,最终调控细胞的生长、生存和分化。代谢通过代谢酶和代谢物直接或间接地调节染色质表达,通过调节染色质反式作用蛋白活性,包括组蛋白修饰酶、染色质重塑复合体和转录调节因子。探索阐明染色质可以被代谢物改变的方式及其功能的重要性仍然是热点研究领域,有助于揭示正常和疾病状态下的基本细胞机制。
参考文献
1. Liu WW, Zheng SQ, Li T, et al. RNA modifications in cellular metabolism: implications for metabolism-targeted therapy and immunotherapy. Signal Transduct Target Ther. 2024, 9(1):70.
2. Dai Z, Ramesh V, Locasale JW. The evolving metabolic landscape of chromatin biology and epigenetics. Nat Rev Genet. 2020, 21(12):737-753.
3. Huang H, Sabari BR, Garcia BA, et al. SnapShot: histone modifications. Cell. 2014, 159(2):458-458.e1.
